The measurement of basilar membrane vibrations in animal species is difficult due to the extremely small amplitude of the vibrations -- less than 0.35 nanometers at the threshold of neural response -- and the inaccessibility of the cochlea, which is deeply embedded in dense temporal bone in many species. To measure cochlear responses that have normal characteristics, the cochlea must be kept in extremely good condition. The sharp tuning that is typical of the healthy cochlea is lost when the cochlea is even slightly damaged. For ethical reasons, direct measurements of basilar membrane motion and single nerve fiber responses are not possible in a living human cochlea.
The lowest level sound that the human ear can detect, at 2 kHz, has a diffuse-field sound pressure of about 20 micropascals. It is customary to use this sound pressure level as a reference when describing the decibel level of acoustic signals; the notation "dB SPL" is used to indicate a sound pressure level in decibels relative to 20 micropascals rms. Under normal diffuse field conditions, this sound pressure level has an acoustic power density close to 1 picowatt per square meter. The acoustic power which enters the ear through the eardrum at the threshold of hearing is very small, about 80 attowatts (1 attowatt = 10^-18 watt). The loudest tolerable sounds are about 120 dB SPL.
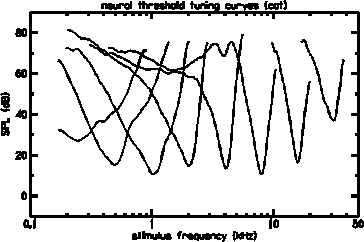
Measurements from single auditory nerve fibers provide only indirect information about cochlear mechanics, but have the distinct advantage that the cochlea need not be opened during the measurement. Neural threshold response tuning curves are an abundant and reliable source of data. Tuning curves, like the ones shown below, indicate the sound pressure level at the eardrum sufficient to elicit a criterion increase in the firing rate of an auditory nerve fiber, as a function of the frequency of a pure tone stimulus. After recording from a nerve fiber, it is possible to mark it chemically so that the path of the fiber can be traced to the place within the cochlea where it synapses (connects) with an inner hair cell (IHC). In this way, reliable frequency-to-place maps can be determined.
Typical neural threshold response tuning curves in cats show a minimum ear canal pressure of about 14 dB SPL at their best frequency and are sensitive to only a narrow band of frequencies. The characteristic frequency (CF) is defined as the frequency with the best sensitivity (or maximum response) for a given place or single neuron. High CF tuning curves tend to have narrower relative bandwidths, with steeper slopes, than low CF tuning curves. The human bandwidths are commonly characterized as being about 1/3 octave.
The first technique that was used successfully to obtain nonlinear basilar membrane frequency response functions in a living (and apparently undamaged) cochlea was the Mossbauer method. This technique measures the Doppler shift in gamma radiation from a small radioactive source placed directly on the basilar membrane. Using the Mossbauer method, Rhode observed that the amplitude of basilar membrane vibrations grew nonlinearly with increasing stimulus level. For example, the figure below shows the level dependence of the ratio of malleus vibration to basilar membrane vibration. If the mechanics of the cochlea were linear, this ratio would be independent of level. Note that the response ratio near CF is largest when the signal level is smallest. This type of input/output relationship is referred to as a compressive nonlinearity. When the animal dies, or when OHCs are damaged, the magnitude of basilar membrane responses becomes smaller near CF and its growth with stimulus level becomes linear.
The degree of similarity between basilar membrane and neural tuning is important because of its relevance to the debate between the resonant tectorial membrane (RTM) and cochlear amplifier (CA) models. It is generally agreed that the tuning measured at the IHC is the same as that measured in an auditory nerve fiber. The IHC tuning curve, however, is not identical to either the isodisplacement or the isovelocity tuning curves for the basilar membrane. The CA models stress the similarities between the basilar membrane and IHC tuning curves and try to account for the observed basilar membrane tuning. The RTM models stress the differences between these two curves and try to account for the ratio of IHC to basilar membrane frequency response. The significance of the differences between basilar membrane and IHC tuning is still a controversial issue.
The ability of OHCs to change their length in response to intracellular voltage changes provides clear evidence of electromechanic transduction within the cochlea and is believed to be the source of the basilar membrane compressive nonlinearity described above. This OHC motility is proportional to the voltage across the cell membrane. The maximum length change observed in isolated OHCs is about 10% of the total length of the cell.
Return to Cochlear Mechanics or proceed to Outer Hair Cell Motility.